
- DEPOD Home
- About DEPOD
- User Manual
- Human Phosphatases
- Protein Substrates
- Non-Protein Substrates
- Pathway Mapping
- Dephosphorylation Network
- ID Mapping

- Download
- Deposit Data
- Acknowledgement
- Contact
- Licence
- Links
- Ensembl
- Entrez Gene
- HGNC
- QuickGO
- Gene Expression Atlas
- OMIM
- UniProt
- IntEnz
- BRENDA
- LocDB
- Catalytic Site Atlas
- InterPro
- Pfam
- CATH
- Gene3D
- PDB
- PDBe
- DrugPort
- ModBase
- SwissModel
- ChEMBL
- Phospho.ELM
- PhosphoSitePlus
- NetwoKIN
- HPRD
- PSICQUIC
- STRING
- IntAct
- MINT
- NCI Nature PID
- KEGG Pathway
- Reactome Pathway
- Pathway Commons
- PubMed
- ArchSchema
- Cytoscape Web
- Europe PMC
- PTP-Central
- EKPD
- HuPHo
- SwissLipids
 Statistics
Statistics
241 human active and 13 inactive phosphatases in total;
194 phosphatases have substrate data;
--------------------------------
336 protein substrates;
83 non-protein substrates;
1215 dephosphorylation interactions;
--------------------------------
299 KEGG pathways;
876 Reactome pathways;
--------------------------------
last update: 11 Mar, 2019
194 phosphatases have substrate data;
--------------------------------
336 protein substrates;
83 non-protein substrates;
1215 dephosphorylation interactions;
--------------------------------
299 KEGG pathways;
876 Reactome pathways;
--------------------------------
last update: 11 Mar, 2019
CHEBI:17202
| Name | IMP | 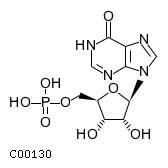 Download: mol | sdf Download: mol | sdf |
| Synonyms | 02'-inosine-5'-monophosphate; 5'-imp; 5'-inosinate; 5'-inosine monophosphate; 5'-inosinic acid; 5'-o-phosphonatoinosine; Imp; Inosine 5'-monophosphate; Inosine 5'-phosphate; Inosine monophosphate; Inosinic acid; | |
| Definition | A purine ribonucleoside 5'-monophosphate having hypoxanthine as the nucleobase. | |
| Molecular Weight (Exact mass) | 348.206 (348.0471) | |
| Molecular Formula | C10H13N4O8P | |
| SMILES | O[C@@H]1[C@@H](COP(O)(O)=O)O[C@H]([C@@H]1O)n1cnc2c1nc[nH]c2=O | |
| InChI | InChI=1S/C10H13N4O8P/c15-6-4(1-21-23(18,19)20)22-10(7(6)16)14-3-13-5-8(14)11-2-12-9(5)17/h2-4,6-7,10,15-16H,1H2,(H,11,12,17)(H2,18,19,20)/t4-,6-,7-,10-/m1/s1 | |
| InChI Key | GRSZFWQUAKGDAV-KQYNXXCUSA-N | |
| Crosslinking annotations | KEGG:C00130 | 3DMET:B01168 | CAS:131-99-7 | ChEBI:17202 | ChEMBL:CHEMBL1207374 | KNApSAcK:C00007224 | NIKKAJI:J9.493E | PDB-CCD:IMP | PubChem:3430 |

| Pathway ID | Pathway Name | Pathway Description (KEGG) |
| map00230 | Purine metabolism | NA |
| map01060 | Biosynthesis of plant secondary metabolites | NA |
| map01065 | Biosynthesis of alkaloids derived from histidine and purine | NA |
| map01070 | Biosynthesis of plant hormones | NA |
| map01100 | Metabolic pathways | NA |
| map01110 | Biosynthesis of secondary metabolites | NA |
| map01130 | Biosynthesis of antibiotics | NA |
| map01523 | Antifolate resistance | Since the 1940s, antifolates have played a pivotal role in drug treatment of malignant, microbial, parasitic and chronic inflammatory diseases. The molecular basis of the anti-proliferative activity of antifolates relies on inhibition of key enzymes in folate metabolism, which results in disruption of purine and thymidylate biosynthesis, inhibition of DNA replication and cell death. The anti-inflammatory properties of antifolate have been most strongly related to its ability to block the release of pro-inflammatory cytokines such as tumour necrosis factor (TNF)-alpha or interleukin (IL)-1beta. Cells may develop resistance to an antifolate drug by virtue of impaired drug transport into cells, augmented drug export, impaired activation of antifolates through polyglutamylation, augmented hydrolysis of antifolate polyglutamates, increased expression and mutation of target enzymes, and the augmentation of cellular tetrahydrofolate-cofactor pools in cells. |
| map04742 | Taste transduction | Five basic tastes are recognized by humans and most other animals - bitter, sweet, sour, salty and umami. In vertebrates, taste stimuli are detected by taste receptor cells (TRCs). At least three distinct cell types are found in mammalian taste buds : type I cells, type II cells, and type III cells. Type I cells express epithelial sodium channel (ENaC) and are considered to be the major mediator of perception of low salt. In type II cells, transduction of bitter, sweet and umami is mediated by a canonical PLC-beta/IP3-signaling cascade, which culminates in the opening of the TRPM5 ion channel. This produces a depolarization that may allow CALMH1 channels to open and release ATP, which serves as a neurotransmitter to activate closely associated nerve afferents expressing P2X2, P2X3 receptors and adjacent type III cells expressing P2Y4 receptors. Type II taste cells also secrete acetylcholine (ACh) that appears to stimulate muscarinic receptors, specifically M3, on the same or neighboring Type II cells. This muscarinic feedback augments taste-evoked release of ATP. In type III cells, sour taste is initiated when protons enter through apically located proton-selective ion channels: polycystic kidney disease 2-like 1 protein (PKD2L1) and polycystic kidney disease 1-like 3 protein (PKD1L3) channels. Weak acids may also activate sour cells by penetrating the cell membrane and leading to closure of resting K+ channels and membrane depolarization. Further, voltage-gated Ca2+ channels are activated and release vesicular serotonin (5-HT), norepinephrine (NE) and gamma-aminobutyric acid (GABA). 5-HT and GABA provide negative paracrine feedback onto receptor cells by activating 5-HT1A and GABAA, GABAB receptors, respectively. 5-HT also functions as a transmitter between presynaptic cells and the sensory afferent. |